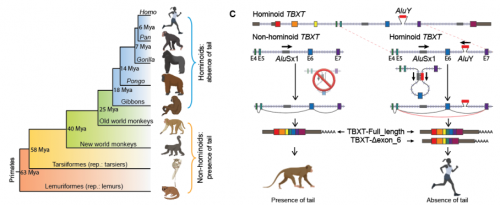
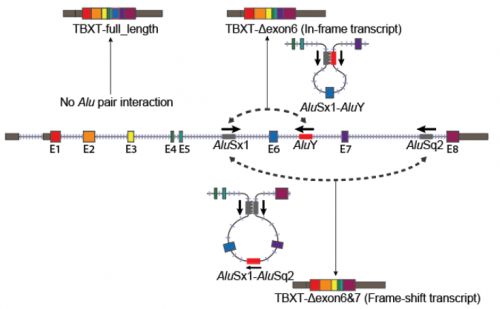
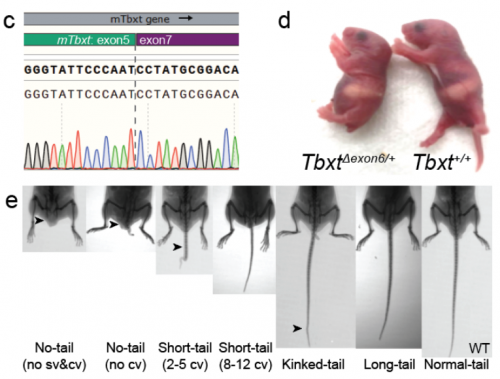
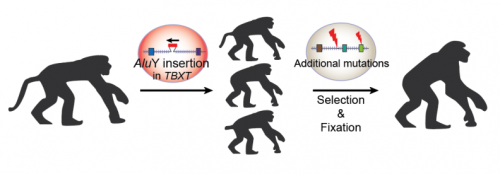
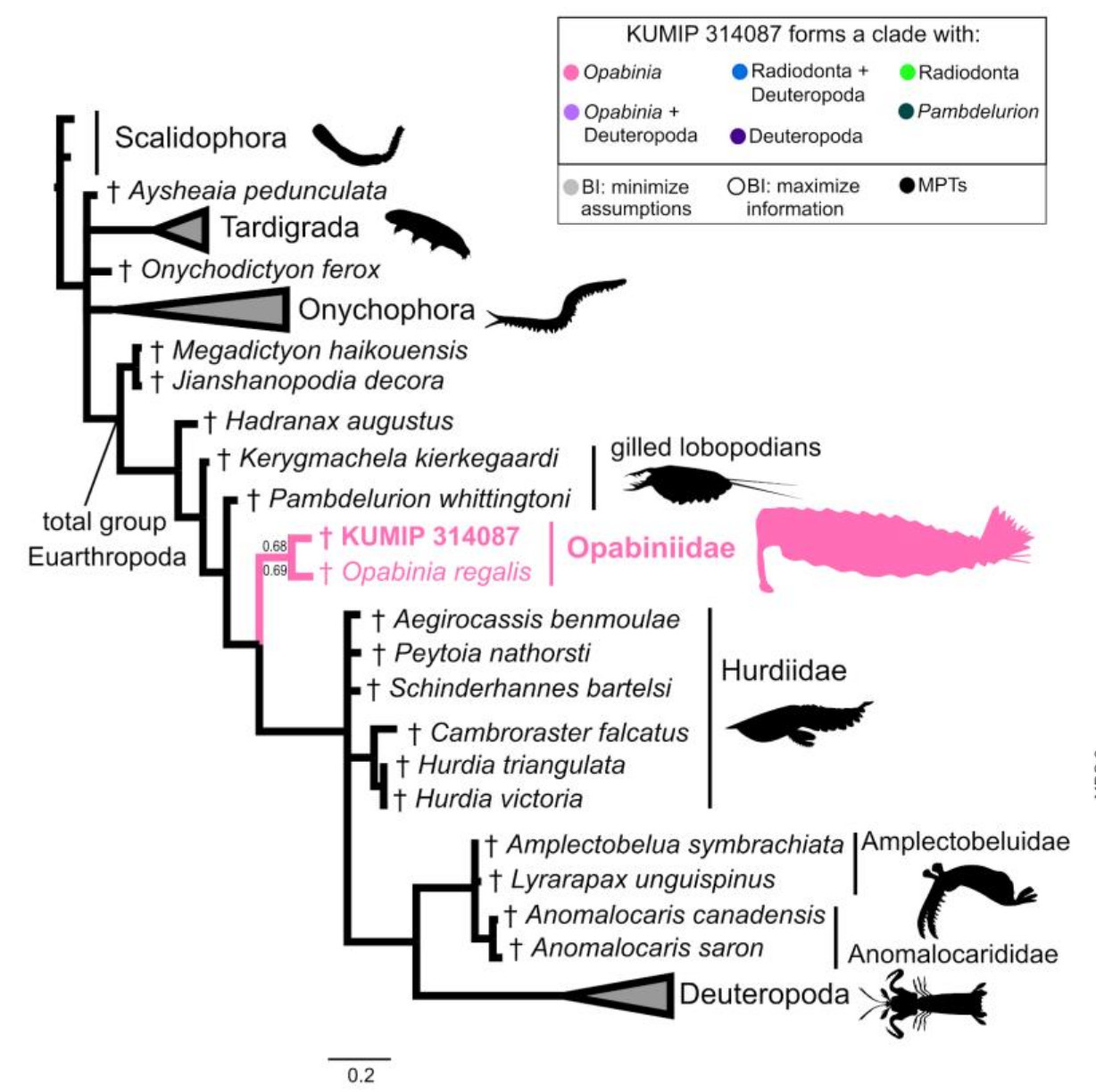
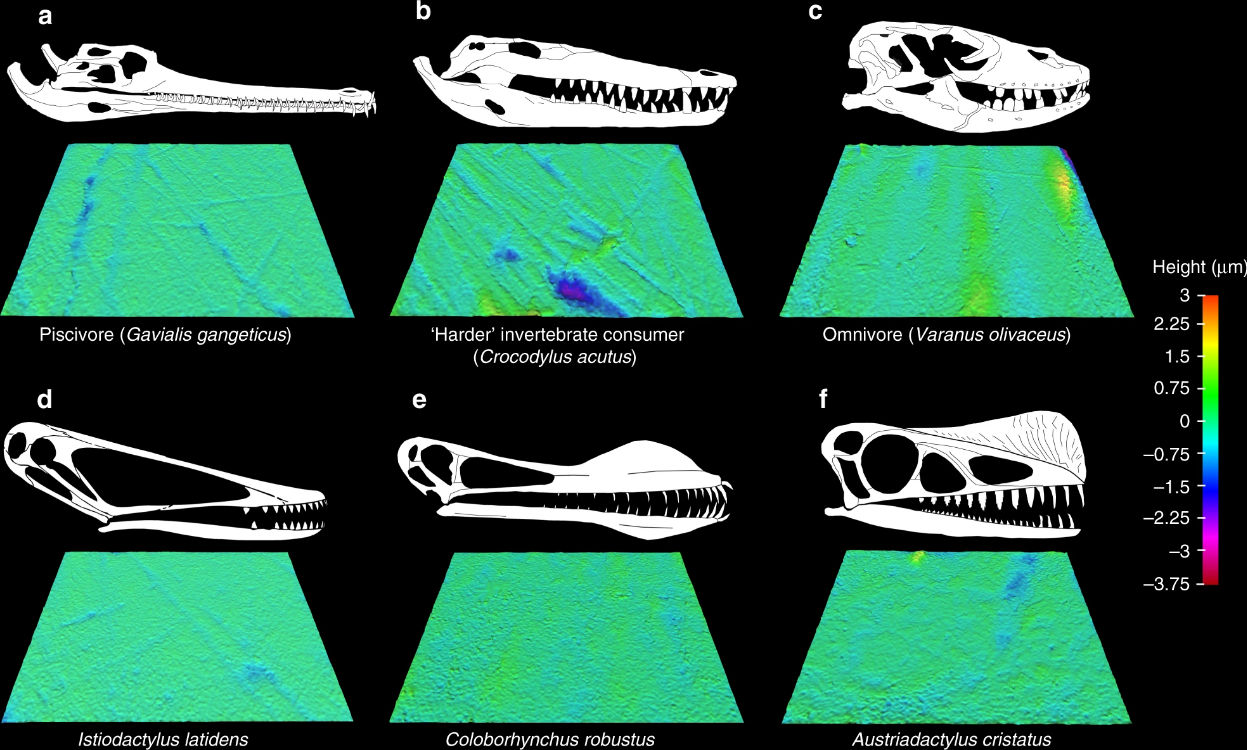
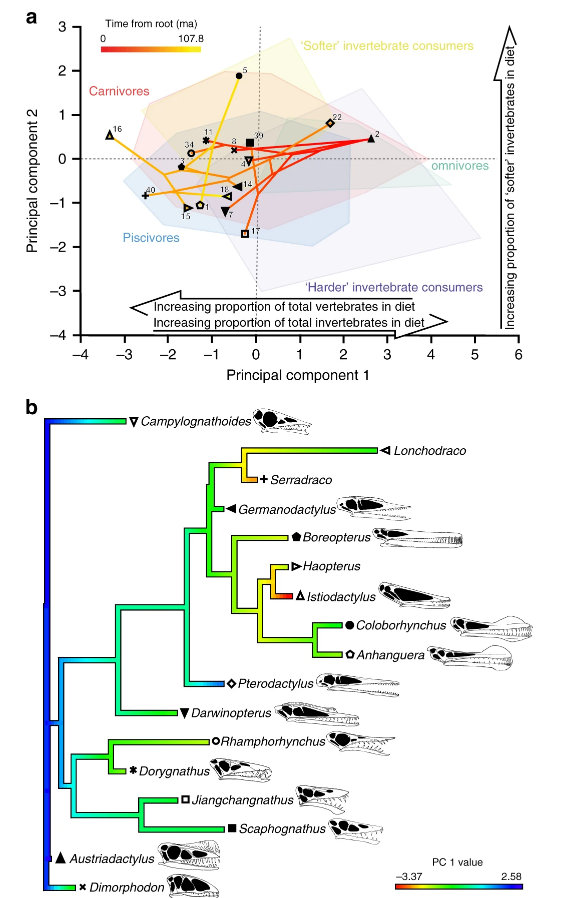
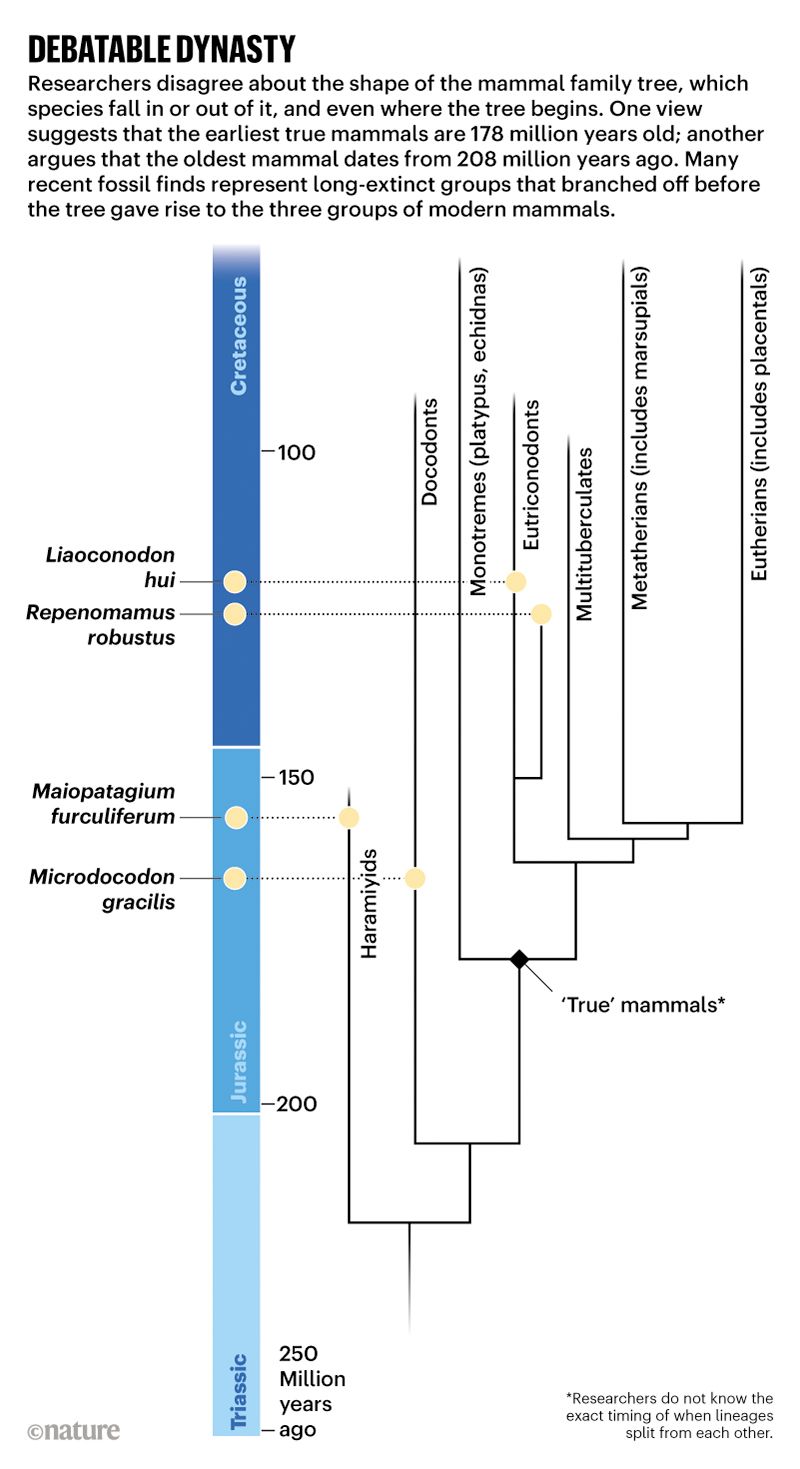
Nice image to illustrate a basic cladistic principle. I still get whines from creationists complaining that I said we humans are fish — but that’s just a bigger circle enclosing everyone in this image.
For the Lisowicia book, number 4 of the Extinct series, we wanted to show the reader how we mammals are related to Lisowicia and other synapsids. It was important for us that the reader understood, in a visual way, that once you are in a group, you never leave the group. pic.twitter.com/6uwMXNGxav
— Gabriel N. U. (@SerpenIllus) October 22, 2021
I have no illusions that this will ever sink into the brains of the people who deny it.