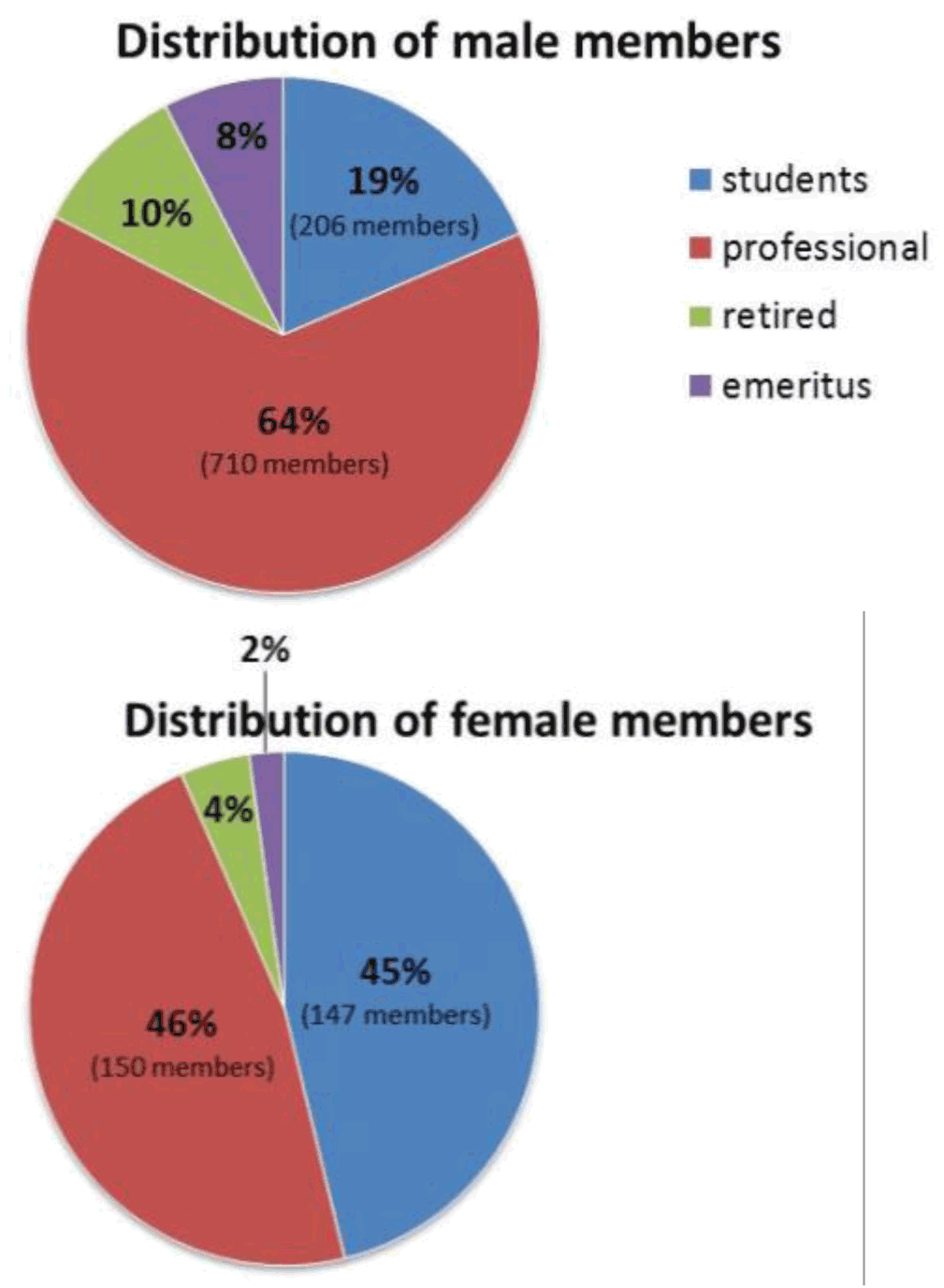
My students are also blogging here:
You know I teach the 8am courses every term, right? Every semester for years I get my oddball classes that weren’t present in the curriculum 13 years ago (when I started here) stuffed into the cracks of the schedule. I’m slowly getting to be a little pushier and am gradually making my way into wakier hours with other classes, but so far, developmental biology is still in the darkness. Fortunately, this talk was so jam-packed with excitement and action that they couldn’t possibly sleep through it! Right?
Just a word about the presentation slides: I’m a firm believer that less is more. My goal is not to display my lecture notes, or lists of bullet point slides that make my points for me, but to show complex and interesting illustrations that I talk about and explain — whoa, I know, how radical. I’ve sat through too many talks that flash 60-80 slides at me in an hour, and it’s too much. Take your time, people! That said, I used 18 slides in a 65 minute lecture today, which I felt was a little excessive — I aspire to someday do a lecture with half that number. But I am weak and need the crutch now.
Also, I returned exams today. People asked if I’d post their answers. No way in hell! These are exams and have the privilege of privacy. I will say that in general the students answered well. The goal of that kind of exam isn’t to confront students with a question that has a specific answer, but with a problem that they should explore, defend, or criticise.
So the subject today was maternal effect genes in Drosophila, specifically the prepatterning information that specifies the anterior-posterior and dorsal-ventral axes. Yes! I can tell you’re all excited!
So I gave them the precursor observations to the actual molecular biology, all this lovely modeling of gradients and information domains that was rich with Turing elegance, and then I dashed their optimism with the cold water of reality: molecular biology has shown that instead of beautifully designed systems, we’ve got bits and pieces cobbled together in a functional kludge. Any pretty patterns we do see are the product of brute force coding.
So they got the overall picture of A/P patterning in flies: a gradient of the Bicoid protein, high in front and low in back, is read by cells to determine their location — its the GPS signal of the early fly. The Nanos protein, also found in a gradient but from back to front, is a hack: it’s only purpose is to clear away a leaky remnant of another gene, Hunchback, which isn’t supposed to be expressed yet (although Nanos may be the diminished rump of a more elaborate ancestral posterior patterning scheme). And the Torso related genes are specifically involved in ‘capping’ the front and back ends of the fly.
The main point of interest about the terminal genes like Torso is their mechanism: we sometimes talk about maternal genes as like a paint-by-number system in which Mom lays out the lines for different areas of differentiation in Baby, and then the embryo fills in the details. The terminal genes are like a perfect example of that: in the follicle, cells literally paint the vitelline membrane of the fly with different informational molecules during the construction of the egg, and then as the embryo develops, these molecules trickle across the perivitelline space (a gap between the outer membrane and embryo proper) to bind receptors and trigger regional differentiation.
It’s also a nice segue into the dorsal/ventral patterning genes, because flies do something similar there: proteins imbedded in discrete regions of the vitelline membrane diffuse to Toll receptors, where they selective activate the Dorsal protein by freeing it from the Cactus inhibitor. We go from a paint-by-number kit to a restored gradient from back to belly side of localization of free Dorsal protein to the cell nucleus. By the way, in case they were getting bored with flies, Dorsal is homologous to NF-κB in us vertebrates, using the same nuclear exclusion/inclusion mechanism, and NF-κB is a hot molecule in biomedicine and cancer research right now.
That was my hour. I closed by threatening them with talk of zygotic genes, specifically the gap genes, next week.
Also, Wednesday we’re going to try something a little different. We’ve finished chapter 5 of Carroll’s book Endless Forms Most Beautiful so they should be ready to weigh the importance of various mechanisms, so I split the class in two and told half of them to read Wray’s article on the importance of cis-regulatory mutations in evolution, and Hoekstra and Coyne’s article that argues for a more balanced emphasis. I’d love to have a fight break out in the room.