I’m on a search committee for a tenure track position in statistics and computer science — we’re looking for someone to teach a data science course, maybe a little bioinformatics on the side, and work with both our statistics and computer science disciplines. I’m the outside member of the committee — you know, the weirdo who isn’t steeped deeply in the culture of the disciplines and maybe is better able to provide the big picture perspective on how candidates will fit with the rest of the university — so I know next to nothing about this stuff. My eyes were crossing and my brain was breaking as I reviewed candidate applications. What I really needed was this bingo card. I think I saw all of those terms fly by as I was flipping through CVs and research and teaching statements.
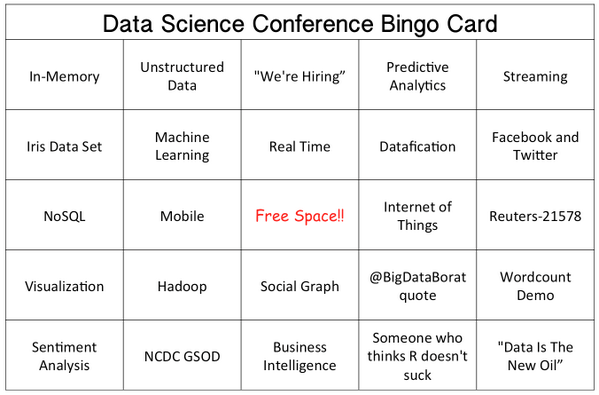
Don’t worry, I deferred to the expertise of my colleagues on all matters dealing with the details of their work.
It’s always interesting, though, to peek into the domains outside my own, and feel a little humbled at all the stuff I don’t know.