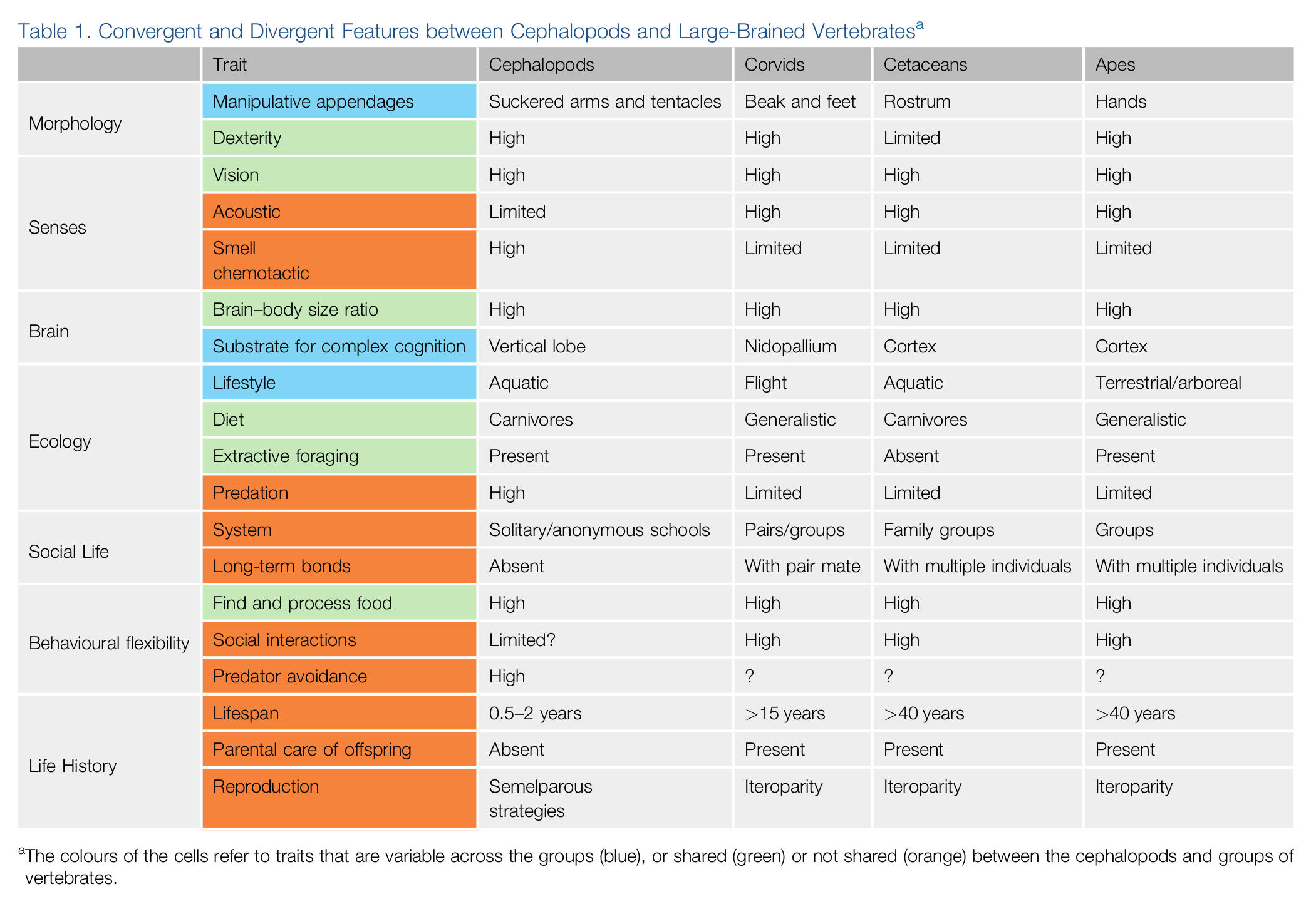
A single piece of paper, handcut to create shading, produces…

That has got to be terrifyingly fragile. Just sweating on it, or handling it roughly, has got to risk damaging the delicate strands of paper holding it together. Put it in a frame under glass, because just looking at it is giving me the heebie-jeebies!