I was rather surprised yesterday to see so much negative reaction to my statement that there’s more to evolution than selection, and that random, not selective, changes dominate our history. It was in the context of what should be taught in our public schools, and I almost bought the line that we can only teach a simplified version of evolution in grade school, but then it sunk in that I was talking to a group of adults about the standard biological perspective, and their reactions were a mix of total bafflement, indignant rejection, and strange evasive waffling. Well, when should we talk about this stuff, then? Do I have to start making day trips to the local nursing home? Or maybe we should be honest from the very beginning about the complexity of modern evolutionary theory and how it has grown to be very different from what Darwin knew.
First thing you have to know: the revolution is over. Neutral and nearly neutral theory won. The neutral theory states that most of the variation found in evolutionary lineages is a product of random genetic drift. Nearly neutral theory is an expansion of that idea that basically says that even slightly advantageous or deleterious mutations will escape selection — they’ll be overwhelmed by effects dependent on population size. This does not in any way imply that selection is unimportant, but only that most molecular differences will not be a product of adaptive, selective changes.
These theories describe different patterns of the distribution of mutations in populations. This diagram from Bromham & Penny (2003) will help you see the difference.
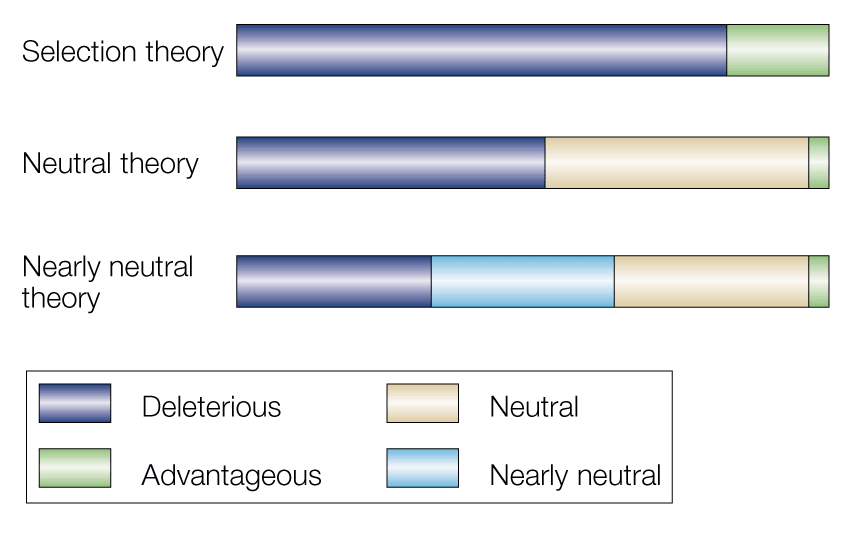
Selectionist, neutral and nearly neutral theories. a | Selectionist theory: early neo-Darwinian theories assumed that all mutations would affect fitness and, therefore, would be advantageous or deleterious, but not neutral. b | Neutral theory: the neutral theory considered that, for most proteins, neutral mutations exceeded those that were advantageous, but that differences in the relative proportions of neutral sites would influence the rate of molecular evolution (that is, more neutral sites would produce a faster overall rate of change). c | Nearly neutral theory: the fate of mutations with only slight positive or negative effect on fitness will depend on how population size affects the outcome.
The purple bars are mutations subject to purifying selection — that is, deleterious mutations that are culled from the population. The green bars are mutations subject to positive selection, that confer some advantage to the individual carrying them. That’s all people thought you would have under old school versions of evolution: every change would have some effect on the individual, and would be subject to selection (and then, of course, smart people started to wonder about genetic load and realizing that there were limitations to how much selection a population could tolerate).
Kimura and Ohta proposed, though, that many mutations would be neutral — that is, changes to the sequence of a gene or the protein would have no effect on fitness, and would be effectively neutral. Since the genetic code is degenerate, with most amino acids coded for by more than one triplet, you could have synonymous changes to the DNA that would produce proteins with identical amino acid sequences. Further, protein structure and function may not be as precisely dependent on specific amino acid sequences as many people assume: there are key regulatory and active sites within proteins that are extremely sensitive to small amino acid changes, but other parts of the protein may be much more fault tolerant. That means that under neutral theory, we have to recognize that beige bar, which are mutations that have no effect on fitness.
Under nearly neutral theory, the domain of selection effects shrunk further, because it was realized that quantitatively, small deleterious and advantageous mutations, that is mutations that only conferred a slight difference in reproductive success, would be invisible against a noisy background of chance variation, and therefore could not be seen by selection. That’s the blue bar; mutations that we can see might cause a slight change to the efficiency of an enzyme, for instance, but are not significant enough to cause any difference in reproductive success, or are either lost or fixed by chance.
Now you might try to salvage your faith in the ultimate power of selection by suggesting that the neutral and nearly neutral mutations are really rare and can thus be ignored as negligible, therefore returning us to the world of selection theory…with just a little fuzzy slop around the boundary between the green and blue bars. That’s untenable, though. We have molecular clocks.
When comparing the rates of change between homologous genes in different species, we had a bit of a surprise: they are very roughly, sloppily constant. That shouldn’t be true under pure selection theory, but it turns out to make a lot of sense under nearly neutral theory. There is a tradeoff in the rate of mutations occurring, and in becoming fixed in a population. A very large population size will accumulate more mutations purely by chance, but the probability of a single mutation becoming fixed in the population is reduced under large population sizes. When you do the math, you discover that population size cancels out, and the frequency of novel forms becoming fixed over time is dependent solely on the mutation rate.
Think about that. If you compare two species, the number of nucleotide differences between them is basically going to be simply the mutation rate times the number of generations separating them from their last common ancestor. That’s how we can use a molecular clock to date the time of divergence of two lineages.
Please note: this does not deny that the selection shapes specific traits in a species occurs — we do undergo evolutionary adaptation! It merely says that most of the genetic changes are random. We have to use specific analysis techniques, like the McDonald/Kreitman test, to detect the signature of selection out of the background noise of mutation.
This is just one example of an important concept that is overlooked when your education in evolution focuses solely on one simplistic version of the mechanisms of change. If you didn’t know it, it’s not your fault; I graduated from high school never having the ‘evolution’ word uttered even once by a teacher, so if you’ve heard about natural selection, you’re one up on me. But we can do better. That the high school level of instruction in evolutionary biology is stuck at around 1930 is a bug, not a feature, and we should aim to improve it.
I know, it’s hard when a significant part of the population is stuck in the first millennium BCE, but we shouldn’t use that as an excuse to dumb down education.
Now, because I so enjoyed the chaos that ensued after rejecting one small part of the Modern Synthesis, let me share with you something rather cool. It’s a table from Eugene Koonin’s The Logic of Chance, in which he summarizes some of the big changes between the Modern Synthesis that emerged in the era before molecular biology, and how most molecular biologists view evolution today. I expect an even more glorious freakout because he refers to this positively as a postmodern reassessment, and I know how much everyone loves post-modernism.
|
Postmodern reassessment of some central propositions of Darwin and Modern Synthesis |
|
|
Proposition |
Postmodern status |
|
The material for evolution is provided primarily by random, heritable variation. |
Only partly true. The repertoire of relevant random changes greatly expanded to include duplication of genes, genome regions, and entire genomes; loss of genes and generally, genetic material; HGT [horizontal gene transfer], including massive gene flux in cases of endosymbiosis; invasion of mobile selfish elements and recruitment of sequences from them; and more. More importantly, (quasi) directed (Lamarckian*) variation is recognized as a major factor of evolution. |
|
Fixation of (rare) beneficial changes by natural selection is the main driving force of evolution. |
Only partly true. Natural (positive) selection is important but is only one of several fundamental factors of evolution and is not quantitatively dominant. Neutral processes combined with purifying selection dominate evolution, and direct effects of environmental cues on the genome ([quasi] Lamarckian phenomena) are important as well. |
|
The variations fixed by natural selection are “infinitesimally small.” Evolution adheres to gradualism. |
False. Even single gene duplications and HGT of single genes are by no means “infinitesimally small,” nor are deletion or acquisition of larger regions, genome rearrangements, whole-genome duplications, and, most dramatically, endosymbiosis. Gradualism is not the principal regime of evolution. [And I would add that even point mutations can have large phenotypic effects. –pzm] |
|
Uniformitarianism: Evolutionary processes have remained largely the same throughout the evolution of life. |
Only partly true. Present-day evolutionary processes were important since the origin of replication. However, major transitions in evolution, such as the origin of eukaryotes, could be brought about by (effectively) unique events such as endosymbiosis, and the earliest stages of evolution (pre-LUCA [last universal common ancestor]) partially relied on distinct processes not involved in subsequent “normal” evolution. |
|
Evolution by natural selection tends to produce increasingly complex adaptive features of organisms, hence progress is a general trend in evolution. |
False. Genome complexity probably evolved as a “genomic syndrome” cause by weak purifying selection in small populations, not as an adaptation. There is no consistent trend toward increasing complexity in evolution, and the notion of evolutionary progress is unwarranted. |
|
The entire evolution of life can be depicted as a single “big tree.” |
False. The discovery of the fundamental contribution of HGT and mobile genetic elements to genome evolution invalidates the TOL concept in its original sense. However, trees remain essential templates to represent evolution of individual genes and many phases of evolution in groups of relatively close organisms. The possibility of salvaging the TOL as a central trend of evolution remains. |
|
All extant cellular life forms descend from very few ancestral forms (and probably one, LUCA). |
True. Comparative genomics leaves no doubt of the common ancestry of cellular life. However, it also yields indications that LUCA(s) might have been very different from modern cells. |
I would add another significant distinction. Under the modern synthesis, populations are primarily seen as plastic and responsive to changes in the environment, producing species that are most strongly marked by adaptive changes. In the postmodern evolutionary view, history dominates — most of the properties of a species are a contingent product of its ancestors’ attributes. “Everything is the way it is because it got that way.” Everything you see in an organism is a consequence of its history, with the addition of a few unique adaptive fillips, and that has two significant implications: you can’t understand an organism without recognizing the impact of its phylogeny, and the modern form preserves ancestral relationships that can be analyzed to discern that history.
And that, of course, demolishes the bogus distinction between historical and observational science that Ken Ham laughably makes. When I observe a fruit fly or a zebrafish or a human being, I am seeing its history made manifest in its structure. See also John Timmer’s recent post on history and science for more.
Bromham L, Penny D (2003) The modern molecular clock. Nat Rev Genet 4(3):216-24.
Koonin EV (2011) The Logic of Chance: The Nature and Origin of Biological Evolution. FT Press.
*I’m not going to get into the evidence for quasi-Lamarckian evolution here, but I will say that there is good evidence for some of it: Koonin discusses the CRISPR-Cas system of adaptive immunity in bacteria. Maybe some other time I can write that up.

I’ll also throw in to everyone not to forget about how what we’re learning about epigenetic inheritance is screwing around with everything we think we know about the modern synthesis. :)
Dear Professor Myers
Is there any way of obtaining this blog post as a PDF?
I found it very instructional, and would like to be able to retain it for future reference even after it “disappears” from view as an older post.
Yours Sincerely
Stephen A Yeats
Very interesting. Des this refined theory have any implications for evo-psychology? Although I’m not an expert in it, I’ve always suspected that the relative lack of accounting for random drift was one of the flaws in many of the evolutionary psychology theories (or, just-so stories). They’re extremely fond of saying “of course trait x was then selected for . . .” without actually supplying evidence that selection occurred in that particular situation.
It’s one reason among many that evo psych is bunk. They are operating from the premise that any difference between individuals reflects an adaptive variation. This is simply not likely to be true.
At some level I understand the argument of “let’s just keep it simple so it’s easier to understand”. But that’s like saying we shouldn’t ask kids to go beyond basic arithmetic in math, or beyond basic sentence structure in composition. No, they can’t learn everything, but they can be told “here’s the most basic idea, but there’s a lot more complexity in how it works out there“. That’s the kind of teaser that can really excite a student into a field.
I haven’t read through this yet, but I have to say I was a little confused by your comments yesterday, so thank you so much for this! Being as I haven’t studied evolution nearly as much as many here, I appreciate the updates from what I learned in high school.
Can somebody simplify this for me (if possible), as in some examples of how these various process might work for, say, different populations of island finches or something?
Hmm, The Logic of Chance sounds like an interesting read!
It is. It’s written primarily from the perspective of viruses/bacteria, though, which will put some people off, and it’s also not exactly light casual beach reading.
Wait.. People have a problem with the “mostly neutral” thing? Do they even think about it at all? I mean, seriously, if all mutations where either bad, or adaptive, then, given the number of bloody mutations in just every single new born, you couldn’t have species, since the accumulated changes would practically make every bloody newborn something completely different from the parents (i.e., damn near the creationist version of having a sheep give birth to a horse). You have to have a large set of neutral/mostly neutral changes, for the genome to remain stable enough to actually have anything like species at all. The alternative is something wildly absurd, like.. X-men style mutant power genetics, where every other random person shoots lighting out of their ass, or has blue skin. I would think that would be bloody obvious.
I am perplexed by your statement about population size: ‘When you do the math, you discover that population size cancels out, and the frequency of novel forms becoming fixed over time is dependent solely on the mutation rate.”
I thought that there was substantial evidence and mathematical support for the idea that small populations (due to isolation or propagating population fronts/migrations) had very much more rapid evolutionary drift; basically from rate going as N and variation as sqrt(N).
Logic of Chance was my first Kindle download when PZ mentioned it was available for free a couple of years ago. I read a little of it from my iPad while waiting in the doctor’s office. I’m just starting on multicellular life. It’s a heavy slog, but it is making sense.
Can you recommend a good online source that provides a beginner-level explanation of evolution as it is currently understood by science? I found this with Google, but is there a better site?
“the high school level of instruction in evolutionary biology is stuck at around 1930” is a bit harsh – I’m pretty sure every high school will mention DNA, after all. Here’s a 1996 paper co-authored by Ohta; she was saying that ‘nearly neutral’ explains things best, but there’s still work to be done to tidy up everything that is known.
I think “stuck around 1980′ would be closer to the truth. I’m not surprised that a lot of us haven’t heard of ‘nearly neutral’ theory before – I can only find one instance of you using the phrase ‘nearly neutral’ on FreeThoughtBlogs – https://proxy.freethought.online/pharyngula/2012/12/11/ep-complexity-is-not-usually-the-product-of-selection/comment-page-1/ – and that wasn’t followed by ‘theory’. If it was theory developed during or after our high school times, and it’s not that frequently talked about, we wouldn’t know it.
doubter – I was about to send you to Berkeley when I realized that was your link. :) That is an extensive and well-done one, and the information in it is solid. You can’t do much better.
So, if I understand correctly, the confusion comes from viewing evolution as “that thing that makes the big visible somatic differences among species” as opposed to the more broad “that thing that changes gene frequencies in populations”. The former presumably is driven substantially by selection, since visible somatic differences are very likely to be exposed to potential selection pressures, but that’s just a tiny subset of what is going on at the gene level, which is dominated by neutral changes, since most nucleotide differences will often be effectively invisible to selection.
If you’re interested in those big visible somatic differences among species, it seems reasonable to say that those effects are dominated by selection both positive and negative. Heck, you could even say that without knowing the precise mechanism of inheritance and mutation. But given that we do know that mechanism, we can see that changes at that level are often not reflected at all in those larger somatic differences.
It’s a bit like assessing the impact of financial transactions at a company by only looking at their mergers and acquisitions, and thus only seeing the huge transactions that have big impacts, and overlooking the day-to-day purchases of paper clips and sales of individual products — those small transactions vastly outweigh in number the huge transaction, but their visible impact may be tiny.
You could also check the companion site to the PBS evolution series here. It also has a lot of good stuff.
1. I do know what was taught in my public schools in a nonfundie area about evolution.
Nothing in 12 grades.
The teachers just ignored it. Except one Mormon teacher in grade 5 who taught us a whole lot of evoution. (This same teacher also used to occasionally preach to us about god in a low key way.)
I”m guessing they were clearly afraid some fundie parents would show up with pitchforks, torches, and rope and didn’t want the hassle.
2. I did learn a lot of odds and ends about evolution though. When I was bored, I would read other sections of the textbook, the ones we never had time to get to,…like the ones on evolution.
I think one source of confusion is different areas of interest. This post is all about genes (which will make Larry happy, I’m sure) — but when the public (even the scientifically-informed and supportive public) hears “evolution”, they tend to think about phenotypical changes — coloration, behaviour, bizarre reproductive strategies (mouth-breeding fish? SRSLY?), size, shape, what-have-you. How much of that is adaptive, and how much “just happened”? Yes, it’s all constrained by history — quadrupeds can’t just sprout functional wings on their backs; the developmental program won’t allow it. And traits are harder to quantify than genes, but they are dependent on genes, in complex ways in which there is rarely if ever a one-to-one mapping between the two.
What we want to know is things like: How and why did some dinosaur lineages turn into flying birds? (Which requires a hell of a lot more change than modifying just scales into feathers and lengthening the forelimb digits). Or some early reptiles into mammals into primates into us?
Genes are important, but there’s more to life than genes.
Thanks, carlie! Reading Pharyngula has made me realize that my understanding of evolution is both fragmentary and out of date. Since I accept evolution as the best explanation for the origin of life, I think I need to understand it better.
I’m going to have to read this several times to really understand it the way I would like to. To see if I’m on the right comprehension track, are these notions essentially correct:
1. that even if the entire environment never, ever changed, a population would still evolve new forms which would eventually be sufficiently different as to be a different species?
2. the original species would likely still continue to exist?
3. the relative sizes of their populations would be… well, I’m not sure how that would work. At this point I realize that my hypothetical spherical, frictionless environment (“environment never changes”) can’t happen, because the organisms themselves are changing the environment by existing it in and mutation?
I’d also suggest that although genetic change is overwhelmingly neutral, it seems likely to me that speciation is largely driven by adaptative changes. So again, the big visible differences in the biological world that people see and try to understand are indeed produced by selection.
@ #2:
I simply copy/pasted the entire thing into Word, made a few tiny tweaks to the formatting (you’ll have to use Word’s formatting to reproduce the table borders), and saved it as a docx file. Agreed that this post will be of great use in discussions with the woo-woo crowd.
BCWebb: Population size completely cancel out for neutral evolution, but is important for nearly neutral evolution. Mutations arise at a particular rate, µ. The total number of mutations in a diploid population, per generation, is 2Nµ, where N is the population size. It happens that the probability of eventual fixation of any allele is equal to its current frequency. And for any particular mutation, that starts as a single copy with a frequency of 1/(2N). Put these together, and the number of mutations fixed per generation comes to (2Nµ)/(2N). The 2N cancels out, and the number turns out to equal the mutation rate.
Then again, nearly neutral mutations actually do have selection working either for or against them. The larger the population, the less important drift is, and the weaker selection can be and still overcome it. So if nearly neutral mutation is important, and if most new mutations are slightly deleterious, those mutations would disappear quickly from a large population but would have a chance of fixation in a small population (at approximately the rate of neutral evolution). If you add in those nearly neutral fixations, a small population is expected to evolve more quickly than a large one.
Tulse: That’s exactly what Jerry Coyne and H. Allen Orr concluded in their book Speciation: most speciation is a byproduct of natural selection acting on geographically isolated populations.
Not necessarily. Generally if all of the traits remain the same and geneflow is unaffected, neutral changes aren’t enough to classify something as a different species than it was before. Humans have lots and lots of genetic differences built up (that’s why DNA forensics works), but not in the traits that “count” for the usual species definitions. As long as geneflow in the population is unimpeded and they don’t develop weirdly different behavioural/physical/metabolic characteristic differences than before, they’d be considered to still be the same species by most.
Shall I assume that the smiley was intended to temper PZ’s wrath? It must have worked, since he didn’t smite you.
What’s the status of the old Dawkins-Gould stoush these days? (I can’t even remember the substance of it. If indeed I ever knew it in the first place).
Things seems very variant and inclusive based on the above there. I suppose those are funny words for it since that’s not what it’s about, it’s about being accurate. But back in the day I can remember people lining up very solidly along specific brands of evolution. I guess they always have though..
I am a female raised in a fundamentalist religous environment – obsequience is my native tongue. ;)
frog: I think there’s some confusion here over what speciation is and what species are. The idea of species as separate gene pools means that speciation is the evolution of reproductive isolation between populations (and in that form works only for sexually reproducing organisms). It isn’t the change, however great, in any single population over time. So your scenario doesn’t have anything to do with speciation; there has to be splitting of populations.
So, to reform your question: could two geographically separated populations, originally identical and in identical environments, become reproductively isolated purely by drift? The answer is that they certainly could and inevitably will, though it would take on average much longer than if there were selection operating on the populations.
I appreciate the effort that went in to this explanation. Unfortunately I never took a college level biology course so while I believe I follow the basic premise of the piece….ie random mutation has a much greater role than natural selection in the evolutionary process, much of the explanation contains so much biological/technical reference and buzzwords I find it very difficult to follow the complete logic. I want to understand, but it’s tough to consider looking up multiple words in every paragraph. I also think some of this explains why a significant percentage of people just accept the simplistic (and silly really) biblical explanation – it’s just easier than trying to wrap around the magnificence of this view of the world.
Yay! I learned something new today! Thanks, PZ!
Back to bed I guess.
Ninja’d by doublereed:
I was here just to say I love being schooled.
I just finished reading “The Violinist’s Thumb” by Sam Kean for a book club and though it’s more about DNA than evolution per se, it does give an accessible explanation of genes and how they work, including some comments about epigenetics at the end. It was published last year so it’s pretty up to date.
I distinctly remember learning that most mutations would have little-to-no effect when I was in high school biology. I don’t remember ever seeing a breakdown of exact percentages or anything, but it was taught here in Utah.
That said, the focus was definitely on the selection part, and probably not just for simplicity’s sake.
Drift is arcane and abstract. Selection is tangible, and it seems likely that selection would be more likely to pique a student’s interest in biology as a whole.
You people that are trying to argue that evolution is somehow different from molecular evolution are breaking my brain. But…but…but…how do you separate them?
Not to say we should be teaching it wrong. It’s not adding a big layer of complexity if you say, “most mutations don’t do anything, but let’s talk about the ones that do something.”
The ‘s’ word seems to me to be the problem through these two threads. If the fundie guy (Mike Fair: see the previous thread) had said “species evolution” instead of “biological evolution” the statement might have seemed a bit less unreasonable: an approximation of the state of play in the time of Darwin (whose book was, after all, called The Origin of Species). But we need to learn to stop being so excited about species. There never has been a consistent definition of species that fits all cases of living organisms anyway, and what PZ is explaining makes clear we need to be less excited about man-made boundaries between phenotypes.
Thanks for the two discussions, anyway, for clarifying our current understanding of molecular genetics and evolution.
@36: Um, because e.g. a limb is not a gene? Even if there is a connection or continuum of some kind between them. And not so much arguing as trying to understand that connection, and what causal processes are at work at various levels.
And you’re not the only whose brain is getting broken this morning ;-).
Re: 4 “It’s one reason among many that evo psych is bunk. They are operating from the premise that any difference between individuals reflects an adaptive variation. This is simply not likely to be true.”
Um… no. They’re using that “premise” to generate hypotheses and predictions, which are then tested. How can someone in your position not understand this? You’re channeling the same nonsense Gould touted about adaptationism 40 years ago! It’s embarrassing, esp. after an excellent post like this about an important topic in evolutionary biology that most layman don’t know about (and should).
frog@21. Essentially #3. Any environment consists of both biotic and abiotic elements. Even if the abiotic environment is static, they biotic environment won’t be because those organisms are mutating too. Probably the closest examples we have for something like that are the experimental evolution experiments with bacterial fermenters that have been going on for years – highly simplified, practically steady state environments, but stuff continues to evolve new capabilities (and pick up genetic load, but evolution is always complicated).
Tulse@22. That’s actually a big area of debate. The null hypothesis for most speciation research is actually the opposite – allopatric speciation is speciation that happens when two populations are separated by some extrinsic factor for a sufficiently long time that they’ve mutated into different species. Most models of adaptive speciation combine some allopatric component with some adaptive component. Examples of actual adaptive speciation (true sympatric speciation) are really hard to prove.
#40, Max:
Two points you seem to have missed:
1. The premise is false. I could generate hypotheses on the idea that the people are descended from elephants, too; it doesn’t make it a useful exercise.
2. They’re tested? Bwahahahahahahahahahahaha!
They are so tested: by comparing them with the gender differences depicted in lifestyle mags, romantic novels and Men Are From Mars…!
Depends on how it’s explained. Some types of drift are easy to understand – give every student in the room a colored bead with about 7 different colors available, have them start milling around, at some point grab two in a corner, quietly have them sit in chairs with their feet up, and yell “OH NO THE FLOOR IS LAVA YOU ARE ALL DEAD” and bingo, genetic variation in the population has dropped from 7 colors to 2, and the only thing those two had going for them was being near you at the right time. It’s a random roll of the dice, absolute unguided chaos, the verification that no, the world is NOT fair, and sometimes there is no good explanation. They might need to avail themselves of a philosophy class after that, though. ;)
I’m reminded of one of the bits out of the Lenski report, where he managed to find an E. coli population that had evolved to eat the citrate base. Specifically, there was a comment that the final mutation that allowed the metabolization of citrate would not have been possible without earlier mutations that made it possible.
The thing is, those earlier mutations only created future potential; by themselves, they were completely neutral. Neutral mutations suddenly became important to selection thousands of generations later… in some strains that got the other mutations.
So ‘neutral’ happens all the time, but sometimes it’s only ‘neutral for now’. Of course, sometimes it’s ‘neutral forever’ because the later changes don’t happen.
This, of course, ties back into the talk PZ gave at Eschaton in Ottawa with regards to the effects of randomness in evolution…
Curses for another learning experience.
I remember some academic saying that my education would have a half life of roughly 5 years. This was back in my college days which is now 8 to 10 half lives ago. This means that whatever I learned then about evolution is now (1-2^(-9))/1 (99.8%) incorrect.
I expect this is pretty accurate but it’s disheartening to have ones nose continually rubbed in the fact. Evolution is not the only subject matter of mine which follows this course.
Thank you though for caring enough to try to fill in.
A humble and grimly surly thanks.
Thanks also to doubter for the link to further, more complete and current, knowledge for me to investigate.
One question for the crowd: It seems that the neutrality of the individual mutations, recombinations, duplications etc is defined only for the current conditions. Wouldn’t the generation of variation itself be an advantageous trait? Doesn’t the label ‘neutral’ for genetic differences presuppose that that point in selection space will always and forever more be unselected positively or negatively? Darwin help me here, but I’m thinking of flex-fuel cars. If you’re stuck on gasoline and the only thing you can get is ethanol you won’t be driving but for now the trait isn’t exceptionally useful.
I KNOW that cars don’t reproduce with variation.
julial@46 There are endless discussions about pre-adaptation, spandrels, blah, blah. What it comes down to is all fitness is contingent. The fitness (positive, negative, or neutral) of any given variant is a function of the physical environment, the biotic environment, the genetic background of the host organism, and it’s frequency in the population – which also means that the fitness of any given mutation is not really best described as an instantaneous value so much as a multidimensional surface. A relatively small number of mutations have a fixed fitness – basically the only ones that are constant are lethal.
Or to put it another way, in the land of the blind, the one-eyed man is king!
…or he’s dead because everyone else suspects him of witchcraft, but hey – life is complicated.
YES. That is another detail that is often left out of teaching about selection – it is only applicable to the current conditions. There are traits that are acted on in some environments but not others, at some points in time but not others. “Neutral” and “selective” are relative terms. However, there are many that are fairly universal. For example, if the mutation results in a human being unable to produce hemoglobin, that will be heavily selected against no matter what the environment. But some mutations will always be neutral no matter the conditions, also. For example, the amino acid leucine is coded for by the base combos CUU, CUC, CUA, and CUG. If there is a mutation there that turns a CUU into a CUC it will show up when you check the bases, but functionally it’s making the same amino acid so would be unnoticeable.
It can be. In fact, the reigning hypothesis about why sexual reproduction exists at all is that it exponentially adds to the amount of genetic variation possible for offspring compared to asexual reproduction.
Yes, and in some cases that may be wrong, but see the above example for some that are. They’re called “synonymous substitutions”. Mutations in noncoding regions of DNA are also almost always neutral.
As a mathematician who works on phylogenetic analysis, I regard selection as an interference with otherwise nice mathematical models, so I’m happy that there’s not much of it about. And since I only got into the field in the last few years, I never had to unlearn the selectionist view.
But I take issue with PZ’s
Go tell that to the trees, because they haven’t heard about this and clearly need a good talking to. See Table 3 from Examining rates and patterns of nucleotide substitution in plants Muse, 2000. It shows that rates of synonymous mutation can vary tenfold (between palms and grasses). Plants don’t leave many fossils so it would be great if they were roughly sloppily clocklike, but 10x is way too much. More strangely, the same table shows that NONsynonymous rates have LESS variation.
@Kagehi (10)
Sigh… Yes, in fact a non-negligible number of biologists have quite a problem with it. I tried to join an evolutionary genetics lab at my alma mater, only to find that they laughed at the entire concept of neutral evolution or.. drift. I had read Kimura’s books by then…
Also, to them the entire genome is functional in case you were wondering.
If you can’t see any negative phenotypes, it’s just because the allele is slightlyyyyyyy negative, if you give the population infinite time and (effective) size any neutral mutation will either disappear or fixate by natural selection. Or something.
@Tulse (16)
Why? This is a bit more complex than it may seem at first sight. Susceptibility to natural selection may not need to be directly related to the “visibility” or any other subjective criterion used to qualify a trait.
Natural selection strongly acts on morphological traits related to reproduction, specially in closely related species (say, cychlid fishes in african lakes [1]). Individuals from these species increase their fitness by being able to sort out potential mates from different species, as interbreeding lowers the fitness of their offspring. These populations’ morphology tends to rapidly diverge under natural selection.
But not all cases are alike. Take for instance human cranial bones, this is a quote from [2]: “Subsequent analyses of temporal bone morphology in modern human populations (Harvati and Weaver, 2006a,b; Smith et al., 2007) have concluded that biological distance matrices constructed using temporal bone shape are highly correlated with neutral molecular distances, while temporal bone size reflects environmental differences related to temperature and latitude.”. Some dimensions vary neutrally, others are shaped by natural selection, and so on.
@Carlie (26)
Hmmm. Actually, there is variation conferring quite surprising traits in most populations and species, probably including humans. But this is treacherous terrain, as this variation normally lies invisible in the form of polygenic traits, thus depending on the genotypes of different loci.
Genotypes do not clearly correspond to phenotypes on a one-to-one basis, and most visible traits seem to depend on the “sum” of the effects of a large number of genes and variants of these. The effect of any outlier variant is completely swamped by the presence of all the other variants, so you won’t be able to distinguish any outlier phenotypes even if the underlying genetic variation is still there.
In the cases where this variation is reduced (say, founder effect, population bottlenecks) or changed (which violates your unimpeded gene flow condition, but there’s no need for these phenotypic differences to appear after the split, they can be lying there, hidden) the variants coding for outlier traits can gain higher frequencies and more prominence in the population.
Some of these traits depend on “thresholds” in order to be expressed, and will only be visible when these are crossed. The existence of previous variation in populations explains the appearance of completely original traits when performing artificial selection on the original populations, such as the phenotypic differences between bulldogs, chihuahuas and wolves. Or how you can artificially select mice behaviors and get a resulting behavior waay above the normal population average [3].
[1] Kerschbaumer et al. (2014) Evolution of body shape in sympatric versus non-sympatric Tropheus populations of Lake Tanganyika
[2] von Cramon-Taubadel, N. et al. (2009) Congruence of individual cranial bone morphology and neutral molecular affinity patterns in modern humans
[3]Gammie, SC: et al. (2006) Artificial selection for increased maternal defense behavior in mice.
Sounds like PZ and I have been thinking about similar things lately! Great post. Anyone interested in neutral theory and contrasting it with the “Modern” Synthesis I would highly recommend reading Arlin Stoltzsfus describing the idea of constructive neutral evolution.
He also wrote a VERY good series of posts on Larry Moran’s Sandwalk about the history of evolutionary theory and how the architects of the MS attempted to combine the findings of the “mutationists” (who often get unfairly painted as misguided saltationists in historical writings about them) with Darwinian ideas about inheritance. The result is the sloppy MS that many people don’t realize modern evolutionary theory is quite far removed from. Great stuff though, he’s a very clear writer and really helped me understand the history of the MS and why neutral theory is such a big deal nowadays. Have you read any of his stuff?
You have to be careful in interpretation, though. These are probably examples not of selection, but of propinquity. History and relationships are real and shape evolution, I think the fundamental problem is when people assume selection must overwhelm them.
But we do have clear examples in molecular genetics of the importance of selection, so we have to be careful not to downplay it. Of course, the most vivid examples I recall from the human genome are the strong signatures of selection in sperm/egg recognition proteins, and in components of the immune system…not the stuff most people want to imagine as being important to their identity as a species.
Also, I was similarly surprised at the intransigence of some professional biologists. As I mentioned, I got no exposure to evolution in high school, and I probably overcompensated in college. There was a fortunately brief period where I was learning evo from popular sources — Robert Ardrey? Lionel Tiger? The early days of evo psych? I has embarrassment — but once I started reading the real stuff, the sound theory coming from rigorous thinkers, I quickly learned what crap that all was. And I assumed that most working evolutionary biologists had been reading Kimura and Ohta along with their Trivers and Hamilton and Maynard Smith and so forth.
But I’ve since learned that many haven’t. You can go through grad school learning how to run a PCR machine and a sequencer and plug data into canned software and get a degree while not learning much of anything. The most recent disappointment was the horrible ENCODE study, which disillusioned me totally.
@ the OP:
You mean most of the variation in nucleotide sequences, specifically.
Yes, it’s such a pain in the ass. If only we could study evolution and selection some different way, like, I don;t know, obsedrving organisms in environments, or something like that. But no, biology has to be about molecular sequences only. A shame.
yay, another book about how Darwin Was Rong, complete with strawmen of what those poor benighted idiots used to think, and lots of special pleading for the importance of the author’s Brand New Hobbyhorses.
Why would you want to do something like that when you could simply sequence its genome? Weird.
I bet he does, and I bet he also has lots of other great examples of the importance of (quasi-) Lamrackian processes, right?
Thanks to LE@47 & carlie@48.
I think I’ll leave the rest of the discussion here to those more capable.
My take:
“It’s Complicated.” (Peter Venkman, Ghostbuster)
re the commnets:
Observe as I deftly ignore the evolutionary-psychology subthread…
oh, bullshit.
tulse @#16&22, Eamon Knight @#19: yes!
hmm, tough one…transcription, translation, and development?
LOL. Yes, that’s why it’s important not to downplay the role of selection in evolution: molecular genetics!
do tell
Yup, with PZ’s recommendation and ChasCPeterson’s anti-recommendation, I’ll certainly be reading Koonin the first chance I get! Unfortunately the Aberdeen U. library doesn’t have it – but it’s not as ludicrously expensive as I expected in papaerback.
True of course–one small hydrophobic amino acid of a protein embedded in a membrane can be the same as another, for example.
But anyone in doubt of the ability of natural seletion to tweak even small, delicate aspects of protein structure ought to read Hochachka and Somero’s Biochemical Adaptation, the book that Larry Moran pretends does not exist.
I haven’t read Koonin, so would not anti-recommend it. I was just going by the contents of that neat-o Table.
@ Carlie (48)
Indeed, in most cases. In fact, different codons, even if coding for the same aminoacid, can be selected for in several cases. For living beings with short life histories such as Drosophila and unlike us, different codons do matter, as the rates of tRNA synthesis are different.
@Julial (46)
Yep, but there’s a paradox here. Say that there is a mutation in a DNA repair gene which causes a higher mutation rate. The ensuing positively selected mutations caused by this increase will be located in different genes, probably located far away from the originally mutated DNA repair gene! Unless this latter is genetically linked to the new advantageous mutations, it won’t rise to fixation alongside them(!).
This really happens in RNA viruses such as Hepatitis C Virus (HCV), where an extremely high muatation rate is selected in order to escape the host’s defenses. Even if more than 80% of produced virions are crippled, the small genome and ultra-rapid selection and expansion of any virions escaping immune destruction lead to a very high mutational load. So high it verges on genomic collapse by mutation, what is called an error catastrophe[1]
[1] Crotty, S. et al. (2001) RNA virus error catastrophe: direct molecular test by using ribavirin.
As a substitute teacher, I use the example of my husband’s hereditary hemochromatosis every time it seems relevant, as an example of just how random evolution can be.
My sources: http://www.mayoclinic.org/diseases-conditions/hemochromatosis/basics/complications/con-20023606
http://www.med.uottawa.ca/sim/data/Biol_Genetics_Hemochromatosis.htm
Hemochromatosis predisposes an individual to many health problems – liver disease, joint problems, heart disease, and diabetes. So, if is so destructive, why is it so common?
“The gene causing the condition is relatively common in people of western European descent, perhaps one in four people have at least one copy of the gene. It might have originated as a mechanism for minimizing iron deficiencies among undernourished people.”
It’s common because it gave survival advantage to individuals with those genes during the many epidemics of Bubonic Plague throughout Europe’s history. The survivors/carriers reproduced, passed on the Hemochromatosis genes to their offspring, and then tended to die young.
(Ironically enough, the best current treatment for the disease is bloodletting. My husband donates blood regularly, to help manage the disease.)
The purpose of all life is to reproduce and pass on its genes. Any gene or trait that does not hinder surviving to reproductive age will get passed on. Random drift.
How do we see this differently if we consider evolution at the level of the organism instead of at the molecular level?
If there a bunch of neutral mutations that result in no change in the production or type of proteins, essentially make exactly the same phenotype (e.g. fall into the redundancy of the genetic code), it strikes me as weird to call these neutral mutations rather than valueless mutations. At the level of the organism, nothing has changed, its like the mutation doesn’t exist.
Because it makes more sense to call a mutation that does result in a phenotypic change – however small – as neutral if it has no net positive or negative effect on the organism.
I guess that is the definition of evolution technically. But I don’t feel like it adds to my understanding of evolution when I’m looking at whole organisms to know that they will experience a bunch of valueless mutations. Actually, it makes it difficult for me to understand how often I should expect phenotype changes to be effectively neutral, because the estimation of the “neutral” category is, to me, artificially inflated with these “valueless” mutations, which, again at the level of an organism, mean absolutely nothing.
In terms of seeing selection as producing everything, I feel like the argument breaks down into confusion based on this: You can’t argue that most of the behavioral differences we see are based on neutral mutations and drift and not selection, when many of the neutral mutations you are citing are valueless and would not produce any differences in the phenotype to begin with (but we are clearly already looking at a difference). It seems logical to me that if we see an actual phenotype difference, it is less likely to be a neutral mutation (as neutral mutations are currently defined). But how much less likely to be neutral I don’t know. Just slightly? Enough to matter? I’ve never gotten a clear picture on that.
16 tulse
22 tulse
I’m missing something. It looks to me (perhaps due to misinterpretation or ignorance) that you’re arguing against the following from the OP, without quite doing so. Which seems to me (repeat parenthetical above) to happen a lot with this subject…
From OP:
19 Eamon Knight
If the area of interest is “big observable change,” then a one can look at the FACT of evolution, and appreciate the big observable change. If the area of interest includes “how do these changes occur,” then it kinda ceases to be a “different area,” and either it’s all the nasty facts or just making crap up. Isn’t it.
And regarding the second part, what does that even mean? That the experience of looking at shape is not synonymous with the process of thinking about process? Again, isn’t that just an appreciation vs. understanding issue?
And isn’t the second part exactly the creationist argument?
what for you have highlighted is the tolerance of life and the environment is to survival. It is not so either or. I do not know if it is cultural or not but we do seem to like to see things as black or white and the closer we actually look the less black and white it becomes.
I can see no great difficulty in teaching the “whole truth” as apposed to some simpler less accurate truth. It would call for saying it differently with a very different emphasis making it very clear that there was or is no goal or outside purpose involved and a lot of “distinctions” have less significance and could be said to be somewhat arbitrary. That kind of makes rote learning teaching more difficult and requires more thinking not, a bad thing.
uncle frogy
#57, Chas:
But who doubts that ability? Not me. The issue isn’t about whether selection can accomplish interesting and powerful things, because we all agree on that; it’s about the ubiquity of selection, and whether we should ignore all the other phenomena that go on in evolution.
53 55 ChasCPeterson
In these posts you appear to be arguing against the OP. But I can’t figure how, since the statements are so, um… terse? Which is none of my business, especially if you mean to be talking directly to PZ (or whoever). But if not, could you make reference to resources on whatever the counter is?
I also don’t neglect it. Another example: work from the Thornton lab on the evolution of steroid receptors. How can you read about that and not realize that you can have remarkable specificity in biochemical processes, but that none of it is determinate or necessary to the course of evolution as a whole?
@ PZ Myers (52)
Indeed! Sorry about my terrible writing, I just wanted to point out that you can’t just straightly rule out the preexistence of genetic variability for phenotypic traits in a population.
@grouperfish (61)
Mutations can change the phenotype of an entire organism and still be neutral.(see the brain bone shapes I described earlier) They’re only neutral relative to fitness, not to the phenotype. And they’re not “valueless”, as already said by carlie, a neutral mutation nowadys can turn into a life-saving mutation tomorrow.
An… Interesting example is a mutation in the CCR5 receptor that effectively reduces the chances of HIV infection. As this mutation is found at low frequency in people of western european descent, a set of hypotheses were presented, namely that this allele protected europeans against the plague and was selected for in medieval ages.
But things ain’t as clear as it first seemed, emphasis mine, [1], [2]: “A much-celebrated story of positive selection in the human genome is the 32-bp deletion in the chemokine receptor CCR5, a variant that confers resistance to AIDS. This variant was postulated to be a relatively recent response to plague or smallpox. New research shows that the frequency of CCR5-Delta32 in Bronze Age samples is similar to that seen today, pushing the observed age of the allele back to at least 3000 and possibly 5000 years ago. Interestingly, the extent of heterozygosity, differentiation across populations and linkage disequilibrium in the CCR5 region is not dissimilar to other human genomic regions, challenging claims of recent positive selection.”
While it confers protection against HIV, a nice phenotypic trait, CCR5-Delta32 doesn’t seem to have been selected in the past, in fact, it looks like it led a rather peaceful neutral life until today. This also calls against facile and fast attributions of positive selection using incomplete population data.
[1]Hedrick, PW. et al. (2011)Using the distribution of the CCR5-Δ32 allele in third-generation Maltese citizens to disprove the Black Death hypothesis.
[2]Hedrick, PW. et al. (2005)”Ground truth” for selection on CCR5-Delta32.
I was taught neutral theory in the last year of highschool! :-) :-) :-)
Unfortunately the pictures are all gone. :-(
Please do, because all evidence for Lamarckian-like evolution I’ve ever seen is 1) the occasional inheritance of histone-level gene regulation for a generation or two, and 2) things like DNA polymerase V (“evolvability”): when things get rough, bacteria switch to more error-prone repair enzymes, increasing their mutation rate – but the mutations are random, not directed as we’d need to reinstate Lamarck.
…But, dude, it was only discovered a few years ago just how many mutations every human has compared to their parents. I mean, for that you need to sequence large parts of three human genomes. That used to take decades! Even sequencing three bacterial genomes for one paper was just about unthinkable 10 years ago!
That was half of the point. They have faster drift – and correspondingly fewer mutations, meaning that fewer mutations are available for drift.
Depends.
1) In the environment in question, how many phenotypically visible traits are under stabilizing selection (meaning that any mutation is deleterious), and how many are neutral or Nearly Neutral™?
2) How do you define “species”? In particular, do you take biochemical traits without influence on the macroscopic phenotype into account?
It goes without saying that all of the above assumes that the species is already adapted to that unchanging environment as well as it ever could be. If it’s not there yet, it’ll change, of course.
If the population splits in 2 (that’s called cladogenesis) and the 2 branches can manage to find a way to avoid competition (by forming separate ecological niches), then yes, depending on your definitions of “original” and “species” (you’re opening a gigantic can of gigantic worms here). If not, the population won’t split, it’ll change as a whole (anagenesis).
…Yeah, pretty much. The organisms around you are part of your environment.
I don’t think that’s true for all species concepts.
No. By definition, nearly neutral mutations are invisible to selection: they have phenotypic effects, but those effects are too small to be selected for or against – drift completely overwhelms selection for or against them. What you’re actually saying seems to be that whether a mutation is nearly neutral or visible to selection depends on population size; and on that I agree.
But a lot fewer than many other generally recognized species! Compared to any subspecies of chimpanzees, we’re all the same.
Raw amount of genetic diversity isn’t a good species concept IMHO…
1) Gould is dead.
2) Punk eek is more common in the fossil record than gradualistic splitting, but both occur – apparently in different environments. This paper (pdf, nice figures) describes the example of a cladogenesis that took a hundred thousand years and happened among diatoms in the entire equatorial photic zone of the Pacific Ocean – the size of that population is, like, huge, man.
3) Dawkins pwnz0rd Gould in Gould’s lifetime on Gould’s puzzlement that so many phyla had evolved in the Cambrian but so few since then: “that’s like looking at an old oak and wondering why it hasn’t grown thick branches lately, only tiny twigs” (paraphrasing from memory).
*long evil laughter*
Depends on your species concept. Yours doesn’t allow for anagenesis as speciation. Several other concepts do.
I approve.
If, on the other hand, it results in some little fish in the Antarctic Ocean being unable to produce hemoglobin, it’ll actually be selected for…
But then, there’s codon usage, briefly mentioned in comment 59: the tRNAs for synonymous codons aren’t made in equal amounts. When there’s less of tRNA for CUU than for CUC, then an mRNA with CUU will be translated more slowly, because it has to wait longer for the right tRNA to end up in the ribosome by Brownian motion. This is a form of gene regulation. If you want to express, say, a human gene in E. coli at full speed, you have to exchange some synonymous codons, because our codon usage is different. (Maybe that should be a TV Trope. ;-) )
O_o
o_O
o_o
O_O
…And that’s just within monocots. Wow. I need to read that paper (which seems to be in open access :-) ).
Well. Sperm/egg recognition contributes to whether or not we can interbreed with chimpanzees* – that’s extremely important to our identity as a species according to the species concept that is by far the most widespread among laypeople!
* It’s never been tried, and there are no good reasons to think it would or would not work…
Well, yeah. That is “most of the variation found in evolutionary lineages”.
Is that an observation or a prediction? …Oh, a prediction, as you explain in comment 58. Bummer.
You know The Alberts? It keeps talking about , meaning all eukaryotes except yeast* – coming up with this moronic term and applying it in this way requires utter and total ignorance of eukaryote phylogeny, which is (funnily enough) known mostly from molecular data.
* Other fungi don’t exist. They’re not model organisms by any stretch, so nobody has heard of them.
Try that sentence again.
This is a semantic distinction. We both agree on the result: drift overwhelms selection if the selection coefficient is small compared to population size (working from memory, s<1/N?; something like that). I, quibbling, would say that there is indeed selection acting on such traits; it's just overwhelmed. And that selection, uneffective as it is, is exactly what separates nearly neutral evolution from neutral evolution.
So, you want to argue species concepts? Speciation concepts? Or perhaps that would be a bad idea.
David M: I disagree with Benton & Pearson on a number of points, most notably about their ability to distinguish vertebrate species in the fossil record and about the relevance of complete geographic sampling.
Fukuda at 67:
If all you have is natural selection, everything looks like an adaptation.
knowknot@62: Sorry to be dense, but I’m not following you. At all. Understand I’ve been kibbitzing on this argument for nigh on 20 years — since PZ and Larry Moran and others would go at it on talk.origins back in the late 90s. And I’ve read my Gould, so I have a nodding acquaintance with exaptation and spandrels and contingency (even if he did overdo it in Wonderful Life). And I still keep feeling like I’m missing something.
Huh? Genes change, by mutation and recombination etc, and some but not all, not even most, of those changes are subject to selection (either because they are truly invisible like synonymous codons, or because the effect on phenotype is minimal and/or irrelevant in the current environment). I think get that part.
But some genetic changes make “big observable change” in phenotype (either singly, or as a cumulative effect). Birds can fly because developmental genes elongate their digits (but only on the forelimbs), grow feathers, grow jumbo pectoral muscles to work the wings, grow a super-efficient respiratory system to keep all that oxygenated, and generally streamline the body and save weight where possible (glossing over obvious diversity among different species, of course). And yet I have a similar suite of developmental genes, and don’t get any of those features.
So when PZ or Larry Moran talk up chance against selection, I think: They can’t really mean all that, can they? I am confused as to what is really being said, and the one time I tried to explain it to someone else, I realized I didn’t understand it myself. So I’d really like, after all this time, to get it straight.
OK, I don’t remember exactly what I was thinking when I wrote that bit, so never mind.
knownot:
Nope, I was trying instead to explain (obviously unclearly) why the average person, including a high school biology teacher, would be more inclined to see “evolution” as being about “the thing that causes all the animals and plants to look different from each other”, rather than “the process that changes gene frequencies”. It’s not that changing gene frequencies, of which most are neutral, isn’t a more accurate picture of evolution, but rather that, precisely because they are neutral, and don’t produce large visible somatic changes, most laypeople would find the origin of morphological difference to be far more interesting than the distribution of neutral SNPs.
It’s the same reason that most medical TV dramas show bizarre illnesses and life-or-death cases, despite the fact that 99% of medicine is people coming in for hangnails and the flu. It’s why police shows focus on murders instead of jaywalking. “Why do different finches have different beaks?” is a far more interesting question to most people than “Why do gene frequencies change over time?”.
And, less flippantly, it’s also why high school physics labs focus on things like phase changes of substances in the rough range of standard temperatures and pressures that most people would encounter in everyday life. You generally won’t learn a lot about plasma physics in high school, despite the fact that a huge quantity of matter in the universe is in plasma form.
Hi Fukuda #67
I’m sorry if i didn’t explain myself very well.
The DNA code for amino acids it redundant. More than one codon (three base pair string) = one amino acid. Often, the third base pair is meaningless and could be anything. So you can have a mutation in a three base codon – especially if it is in the third position – that leads to the “calling” of the same amino acid. So you can have a change in DNA that does not create any single change in the creation of a protein. There is no way it can effect anything. The mechanisms for reading DNA will read it exactly the same way as before. It’s less than neutral, it’s valueless.
I am arguing that these types of mutation should be thought of differently than true neutral mutations when result in phenotypic change that is neutral in the face of natural selection.
I get confused when people talk about the prevalence of neutral mutations when they include “valueless” mutations in the mix.
grouperfish,
You want to change the definition of a term that’s commonly used in biology, i.e. “neutral”. And you are welcome to try, though it will prove difficult. But I don’t see why you should be confused when other people use the term in its standard definition. Most neutral mutations have no effect on phenotype. Deal with it.
By the way, most neutral mutations (at least in eukaryotes) are not in protein-coding regions of the genome; they’re in that portion of the non-coding regions we call junk DNA. (Necessary clarification: “non-coding” and “junk” are not synonyms.) And it’s not necessarily true that silent substitutions in protein-coding genes are not subject to selection; as others have mentioned, there may be weak selection favoring certain synonymous codons, especially in genes or taxa with high rates of translation.
This shocked (and infurated(s)) me when I first encountered it too. There is currently a (minor, amicable, and mostly joking) feud between my former undergrad advisor (who teaches evolution and ecology) and another prof (who teaches genetics) about whether neutral mutations actually exist and whether nearly neutral mutations actually have meaningful selection acting on them. I know the geneticist understands effective population size, so I don’t understand his intransigence.
Even more shocking to me was when I did my undergrad thesis. Speaking vaguely, because I’m still polishing it for publication and its something that could be replicated in a short while by a full-time researcher with full-time access to a supercomputer, there’s some weirdness in the molecular clock which is commonly ascribed to selection on nearly neutral mutations. In primates, who universally have low population sizes. My work basically showed that it can be explained in its entirety by drift and/or the interaction between uncertainty and biases in estimation, but to the best of my ability to find out, nobody’s bothered to look at this possibility.
Another total amateur here, so please moderate hurled objects accordingly.
Koonin: There is no consistent trend toward increasing complexity in evolution…
I can’t help but rate that as “partly true”. Consider, for a vague phenotype-level example, a hypothetical eight-legged creepy-crawly way back when. Some individuals within that population discover that certain bodily secretions can, say, trap smaller creepy-crawlies, which comes in handy for nutritional purposes.
Three million and five generations later, our proto-spiders are leaping from leaf to leaf, oozing their goo in strands behind them and thereby trapping some flying bugs too – yum! Seven million and seventeen generations later, the hot new fad is webs, man – if you can’t make elaborate radial & concentric formations, you won’t even ever get a date.
And so on and so forth, just to produce a rough example of “increasing complexity”.
None of this means that, say, amoebas, can’t keep doing their thing without finding any use for either a bell or a whistle. Still, complexity has increased along that one branch of organisms, in ways absolutely dependent on accumulation of previous increases. We also see parallel developments in other populations, such as some of those tasty bugs going through multiple metamorphoses in a single lifespan.
So – setting aside Chicxulub-type resets and the like – I do perceive evolution consistently producing increasing complexity, even if some organisms (e.g., E. coli, sharks) have found a sweet spot where added frills only get in the way.
Where have I got it wrong, Prof. Koonin?
OK.
Quite the opposite. :-) I’m anxious to make sure everyone keeps in mind that “species” means different – overlapping to contradictory – things to different people, and not rarely several different things to the same person. I don’t mean to single you out.
Ditch the word “species” and replace it by “distinguishable phenotypes”, which is what they’re talking about.
Oh yes. That’s a major, major issue in the continental fossil record.
The difference between you and a bat is… basically a bit of gene regulation, which is heritable because it (AFAIK) consists mostly of mutations to promoters, enhancers, terminators, silencers and whatnots.
@78: I don’t know about Koonin, but you’ve more or less got what Gould said in Full House. He argues that, given life started out maximally simple, there was no where to go but up. However, that does not mean there is a universal tendency for any given lineage to add whistles and bells over time — most life, in fact, is still happily prokaryotic. Thus “life has gotten more complex over time” is true only as a statement about the upper limit achieved, across the entire biota of the planet, not about individual lineages. (It may, of course, still apply about one’s own life ;-)).
Lots of places.
1) You’ve confused increasing complexity with increasing diversity of complexity. Also, keep in mind that complexity can’t decrease below a minimum, because below that minimum life isn’t possible, while the other end is open.
2) It’s not “some organisms”, it’s the vast majority of all life that’s not under selection for increased complexity. Indeed, many bacteria are under selection for faster reproduction and therefore smaller genome size.
3) Parasites, even multicellular ones, are under selection for faster reproduction at the expense of pretty much everything else. They are under selection for getting less complex. There is no comparable guild that is under selection for getting more complex.
4) Many amoebas have mind-boggling amounts of junk DNA. :-) Most bacteria have none.
5) Sexual selection (“won’t even ever get a date”) is always in compromise with natural selection. That means that wherever it doesn’t change in direction or intensity all the time, it leads to extinction.
Just off the top of my head.
(Yes, Gould’s Full House is the go-to book on this topic; I’ve taken most of my arguments from there.)
Do we even know that last part for sure? Could there be some theoretical upper bound for complexity, above which there are simply too many interacting parts, and the whole thing is simply prone to fall apart spontaneously?
I think an argument could also be made that the overall complexity of life on earth has not changed since at least the Permian, if not earlier. I mean, putting human beings aside (so we can ignore any chauvinistic thoughts of the “special” complexity of the human brain), can we really say that even the most complex modern organisms are truly more complex than the most complex of ancient ones? Is a wolf more complex than Dimetrodon? Is a scorpion more complex than Anomalocaris? Mammals more complex than dinosaurs?
?
There’s no reason at all why sexual selection pressures (i.e. those that promote mating success) can’t push in the same direction as selection for other components of fitness. In fact, that concept is at the core of the various “good genes” hypotheses of sexual selection.
raefn@60:
As someone with a family history of hemochromatosis (and its treatment; my grandfather was getting a pint of blood taken out of him every week for a while, and the hemochromatosis caused his diabetes), I had thought that part of the reason it was so prevalent was because the higher blood iron levels were potentially an advantage to women of early child-bearing years, who can otherwise often suffer from anemia. Hemochromatosis is one of those things that rarely seems to kill anybody until they’re well into their forties anyway, at worst, so it’s not really strongly selected against.
On the other hand, your Bubonic Plague mention would definitely explain why it’s far more common in people with primarily European genetic history.
I really need to read The Logic Of Chance, it’s been sitting on my kindle app for far too long. That and Making Sense Of Evolution will be the next books on evolution I read.
David,
Well, sort of. If you use a strict morphospecies concept, then punk eek reduces to a claim that the evolution of distinguishable morphotypes occurs mostly during episodes of the evolution of distinguishable morphotypes (what speciation means under that concept), which is not as interesting as Ann Elk’s theory about the Brontosaurus. On the other hand, if by punk eek Benton & Pearce mean merely that morphological evolution common occurs in episodes, not uniformly over all time, then that’s also not as interesting but in a different way.
I’ve found this website to be quite useful for my high school bio students, in both its breadth and depth.
http://phylointelligence.com/evidence.html
@Amphiox (83)
McShea[1] actually bothered to quantify the cellular complexity of unicellular vs multicellular prokaryotes, actually confirming that individual cells within a multicellular organ lost much of their original complexity while the overall organism increased in complexity (about the hypothesis):
“Concretely, the hypothesis predicts that the cells of multicellular organisms will have fewer part types than free-living eukaryotic cells, that is, nonparasitic protists (hereafter
simply protists; sensu Patterson 1999). At a higher level, in colonial marine invertebrates and social insects, the multicellular individuals (e.g., polyps, zooids) that constitute the more individuated colonies—that is, those in which coloniality is more highly developed (Boardman and Cheetham 1973)—should have fewer part types, on average, than those of their less colonial or solitary relatives (Beklemishev 1969). And at a lower level, the former prokaryotes that today function as organelles in eukaryotic cells (e.g., mitochondria, plastids) should have fewer part types than free-living prokaryotes.” At a “higher” level, colonial bee brains for instance are noticeably smaller than the brains from other related non-colonial bee species.
Also, about the human brain… I have bad news for the chauvinists [2].
[1]McShea, DW. (2002) A complexity drain on cells in the evolution of multicellularity.
[2]Herculano-Houzel, S. (2012) The remarkable, yet not extraordinary, human brain as a scaled-up primate brain and its associated cost.
David Marjanović @ # 81 – I had thought my contrived example assisted my argument, but it seems to have had the opposite effect. Let me try again more generally.
Biological evolution creates opportunities for increased complexity because it works accumulatively; some of those opportunities expand into successful niches, and allow further opportunities and compounding complexity.
Though, as you note, such elaborations often fail, for very good reasons, enough succeed that a time traveler seeking highly complex organisms would find more of them (as species, not individuals) in later parts of periods rather than earlier (“period” here meaning interval-between-mass-extinctions).
My case suffers from lack of a good definition of “complexity”, among other things – one reason why I included my fanciful spider-evolution story to attempt to illustrate by example. And I limit my case to phenotypes, not knowing enough genetics to even dip my toe in those waters.
Though bacteria and parasites have good reason to K.I.S.S.,* and highly complex organisms have regularly specialized themselves into extinction, the overall evolutionary process, as partially seen on this one planet, does produce a subset of increasing complexity.
Doesn’t it?
*Keep It Simple, Sweetie
Wow. As a decently educated layman, I thought I knew the rudiments of evolutionary theory. Oh dear. This is the first time I have ever even heard of neutral theory and nearly neutral theory. My knowledge is 20 years out of date. I read the article once, then a second time, then a third, and was finally able to get the gist. Thank you so much. Oh, and thanks, too, to Carlie. Are you also a professional biologist? Oh, and bryanfeir, the lightbulb lit up when you explained how “neutral” could mean “neutral for now,” but not necessarily in perpetuity. I will try to keep up, but probably not in great depth, as I am an old lady at the outer fringes of circling the drain and have yet to taste other aspects of life, while being constrained by the limitations of being a caregiver. But I’m dancing just as fast as I can.
I’m basically with “tulse”. It is worthwhile to keep clear the distinction between phenotypic and genotypic evolution. The stereotypical characterization of the modern synthesis applies more to the former than to the latter (and it is likely to be the former that is most of interest to the lay public). This neo-Darwinian, for one, is quite comfortable with genome level changes that are not “infinitesimal”. I argue only that large changes in phenotype are seldom favored by natural selection. I still like Fisher’s classic analogy to a slightly out of focus microscope: a small change has, at the limit, an almost 50% chance of being an improvement; a large change will almost always make things worse. But, again, we are talking about phenotype, not genotype. Some “large” changes in genotype (e.g., duplication of a gene for which presence of function is dominant to absence) will have small or negligible immediate phenotypic consequences. Now for a pet hypothesis: speciation often depends on a combination of drift and selection: during population bottlenecks, nearly neutral (but slightly deleterious) mutations drift to fixation. As the population expands, select takes over and favors some “fix” for those deleterious effects, But often, these are “second site” mutations that, by chance, differ from one population isolate to another. Put such populations back together, and the differing sets of problems and fixes will not work well together, causing selection against hybrids. If I weren’t happily retired, I might try to work that idea up for publication. Maybe somebody already has?
leepicton – there are a lot of biologists on the thread! That’s part of why you’re seeing so much fighting – sitting around and yelling about theory particulars is normal operating procedure. :)
That’s what it looks like to me.
David Marjanović went
From Cho et al:
Cho Y, Mower JP, Qiu YL, Palmer JD. 2004. Mitochondrial
substitution rates are extraordinarily elevated and variable in
a genus of flowering plants. Proc Natl Acad Sci USA.
101:17741–17746.
Also:
Parkinson CL, Mower JP, Qiu YL, Shirk AJ, Song K,
Young ND, DePamphilis CW, Palmer JD. 2005. Multiple
major increases and decreases in mitochondrial substitution
rates in the plant family Geraniaceae. BMC Evol Biol. 5:73.
and:
Daniel B. Sloan, Camille M. Barr, Matthew S. Olson, Stephen R. Keller,
and Douglas R. Taylor
Evolutionary Rate Variation at Multiple Levels of Biological Organization
in Plant Mitochondrial DNA Mol Biol Evol (2008) 25 (2): 243-246
ah, mitochodrial DNA. You didn’t say that the first time. Still a surprising range of variation, but not as weird as it would be for the nuclear genome.
Can someone please answer the following stupid question, which comes from my own personal, uninformed head, in the simplest terms possible?
|
IF there is a FACT of evolution, which is made evident by demonstrable physical change extending from obvious physical traits down tomthe level of the genome
AND IF it is commonly understood that the MECHANISMS of evolution (with which THEORIES of evolution are concerned) are acknowledged to be changes in the genome due to mutations and effects of the environment (including all the subtle possible interactions these imply)
AND IF any individual form and function depends ultimately on restrictions and possibilities made present in the genome
|
THEN how is it that, even though physical change can be appreciated strictly in terms of the changes themselves, any meaningful study of the mechanisms of evolution can be undertaken without specific and thorough reference to the genome itself?
|
PLEASE, PLEASE, PLEASE understand that I’m not being silly, or caustic, or ironic here, nor that I think I know a DAMN THING.
|
I’m not surprised that I don’t yet get the intricacies of this argument, and I expect very little outside help with that. But I AM surprised that I can’t comprehend even its SHAPE.
@Pierce R Butler
What Koonin is saying here, I think, is that if you’re looking at your hypothetical protospider through your time machine, there’s no reason to predict that it will evolve a complex web-spinning system for catching tasty bugs rather than remain a protospider or even become a parasitic mite and lose the capacity for free-living entirely. Sure, over time some taxa become more “complex”, but that’s not true when taking all organisms into consideration.
Uh, the reason that evo psych is almost always bunk is that it attempted to say A = B, or is derived from B, as the case may be, then, when asked to do the math, at some point in the process, sometimes right at step #1, tried to divide by zero. I.e., they are trying to claim that 3 is derived from 2, by adding X, which can be any number from 1, which might make sense, to 34,454,663,534,352,545, which doesn’t, by simply misplacing a whole mess of math from the middle of the process, or, substituting (current_result)/0 + 2 + (some other stuff) = 3.
It all a game of, “Some where in between, in that window of fuzziness we can’t describe, explain, or even make tangible claims about, something happened, which allows 2 = 3. Even if they where right, it would be by pure bloody accident, since they can’t show any verifiable connection between the behavior, and their premised “cause(s)”.
Umm. I seem to remember reading something some time back about there being some problems trying to engineer variations of some plants, because they had a more than usually robust cellular repair system in place, which would result, when confronted with a broken gene, in them substituting, so long as a second unbroken copy was available, the unchanged version, such that, eventually, all new cells in their growth cycle contained only the repaired genetic patterns, and none of the artificially introduced edit. I can certainly see the possibility of, perhaps, some plants having adapted a less precise repair system too, producing a wide variation in mutation rates, due to, in one, nearly all changes being erased by repair mechanism, and in others, almost all changes retained, due to few, if any, such repairs being done at all.
Well, one solution to that problem would be to turn off most of the parts, if not needed, specialize the parts that are turned on, and diversify the cell types, to get this result, and, while at it, limit the level of interaction between these various parts, such that chemicals not needed for cell type A and not being taken in, and reacted to, but are, for type B. I think there is some sort of special name for organisms like this… lol
But, seriously, there may be some upper limit, but its hard to say just what it might be, though.. it might be a reason why symbiosis is easier to manage than having “every single” function needed to, say, digest food efficiently, in place, all in one organism, is uncommon/non-existent in complex forms.
Of course there could be – Gould’s point is that there’s no reason to think any lineage has ever approached it. I should have made this explicit.
We probably can’t say any such thing. Complexity is hard to define, though, or rather the definitions that make sense tend to be hard to apply.
Well, there’s this loop in the “honest signal of fitness” hypothesis… in such cases, sexual selection selects for wasteful, harmful things against all selection for efficiency, but by selecting for the ability to maintain all this counterproductive crap, it selects for efficient and/or effective use of energy from the environment more strongly than natural selection alone would do. :-)
That’s what it means.
Even this is less true than everyone used to think. Today’s alligators are descended from a long line of specialized crushers with rounded, almost hemispherical, teeth in the back halves of their jaws.
:-}
45 years in the case of neutral theory – Motoo Kimura published in 1968.
To my own astonishment, nearly neutral theory is 40 years old: Tomoko Ohta published in 1973! I was minus nine years old!
Since she declined to answer… yes :-)
I think you’re talking about handicap hypotheses. Other versions of honest signal/good genes do not require anything wasteful or harmful. For example, bright red carotenoid coloration may be an honest signal of both food-gathering ability and immune-system/antioxidant-related health.
And then there’s intrasexual selection: being big and strong enough to win fights is also big and strong enough to ward off predators; being fast enough to defend a large territory is also fast enough to catch a lot of bugs, etc.
My father’s self-deprecating version: Keep It Simple, Stupid Swede (yes, both his parents were Swedes; no, he’s not too stupid)
That reminds me, Paul Nelson Day isn’t that far off. For those who don’t know, the day is named after an intelligent design creationist who attempted to quantify complexity.
It was really bad attempt. There are much better measures of complexity. Since some of these are uncomputable even in theory, applying them would indeed be very difficult.
I don’t think we can be sure that endosymbiosis (as in the origin of organelles) is not gradual.
I occasionally quote some research that if a trait has 2x% adaptive advantage, it has x% chance of becoming established in the population and 100-x% chance of being lost; and I get polite incredulity.
Note that this calculation is based on an initial frequency of 1/(2N).
Nepenthe @ # 98 – thanks for a helpful reply.
I’m coming to think that Koonin was simply denying a widespread misunderstanding of “evolution = progress” which leads many people to think that increased complexity is basically inevitable across the board in all organisms.
Whereas I was looking only at the production of complexity anywhere, and seeing that it does increase in certain particular (I almost said “specified”…) cases.
@97 knowknot
I’ve read this over a couple of times and I can’t really parse the question; perhaps that’s why no one has answered. Rephrase?
@Pierce R Butler
Pretty much.
And credit to Eamon Knight @80, who’s post I did not read, but was earlier and more concisely stated than mine.
I think Koonin’s table is a good start, but I would suggest several revisions.
The first one is about “the material for evolution”. Not only do we know that mutations are not all infinitesimal and not (by various rational scientific definitions) “random”, we no longer believe that they are “the material for evolution.” Here is an exercise that I found very fun: do a google image search on “raw material” and study the results. Now ask yourself the SAT-style question, is the relationship of mutational inputs to evolutionary outcomes like the relationship of raw materials to factory products? Dobzhansky used that metaphor of the raw materials for a factory, because his theory of evolution said that variation always comes in masses of individually unimportant effects. He literally said that an individual mutation is unimportant, like the movement of an individual molecule in a gas, and that the causes of evolution are like the gas laws, in that they happen at a higher “population” level. This is a metaphysical theory about what kinds of things are causes, but it is also a ridiculously narrow theory of population genetics. Either way, it’s something we need to get away from. When evo-devo people talk about “the arrival of the fittest” they are offering an alternative metaphor to “materials” (no one in evo-devo really believes in “raw materials” but the phrase occurs surprisingly often in the evo-devo literature). Referring to “material” rather than “raw material” is an improvement, but it isn’t enough. Part of the issue is that variations incorporated in evolution are not all infinitesimal (which Koonin covers), but it is also that they are dynamically a source of initiative and (at least metaphorically) a source of creativity. The “materials” metaphor goes hand-in-glove with the religulous metaphor of selection as the potter (or other artist or craftsman) that shapes the clay.
The second one that I would change is the proposition that selection of rare beneficial mutations is the driving force of evolution. I can see what he is getting at in the list of alternatives, but the orthodox proposition is mis-stated. The architects of the Modern Synthesis said very clearly that selection does not wait around for rare beneficial mutations. They thought of this as mutationist or mutation-driven evolution, and it is definitely a post-synthesis (and pre-synthesis) view. The Modern Synthesis view was that there is always abundant variation in the “gene pool” for selection to craft an exquisite adaptation when the environment changes (again, picture selection as the artist, like a painter with a palette full of pigments). Like the above, this is both an ideology about causation and a theory of population genetics.
So, I would re-state this proposition, and I would stress what I think is one of the most profound and well evidenced changes in evolutionary thinking, which is that we now accept many aspects of mutationist thinking that the architects of the Modern Synthesis rejected in their quest for a more ideologically pure Darwinian theory. Darwin literally said that a theory of the acceptance or rejection of individual true-breeding mutants “may be a very good theory, but it is not mine”. His assumption was that evolution is always about shifting masses of small variations, and the Modern Synthesis followed dutifully along with Darwin. Today we have keynote speakers like Allen Orr and Dan Weinreich who present models of adaptation based on a mutationist origin-fixation process. They call this “Darwinian adaptation” but they ought to call it Jenkinian or deVriesian adaptation because Darwin and his followers rejected it.
I also don’t think the idea of a single large tree is characteristically Darwinian. Does he think that Lamarckians believed any differently? This is really confusing mechanistic theories about evolutionary causes with common heuristic assumptions. If the scientific world agrees on theory X and also assumes Y, then everyone who believes in X also believes in Y. But this does not mean that proposition Y is part of theory X. Every architect of the Modern Synthesis ignored lateral transfer along with plate tectonics, mobile elements, prions, etc. Lateral transfers are contrary to Darwinism, not because they screw up phylogenies, but because they signal the real evolutionary importance of macromutations, creative change-initiating events that can’t be squeezed into the “raw materials” metaphor.
I also don’t think the idea of a single large tree is characteristically Darwinian. Does he think that Lamarckians believed any differently?
I don’t know about Lamarckians (were there ever any?), but as far as I can tell Lamarck believed in many separate origins of life all through earth history, with each of them following a modified (by being just a bit branched) scala naturae. Darwin may have been the first to consider life to be a single tree (though he waffled on that and allowed for a small number of separate trees).
If I’m understanding you correctly, it’s the size of the effects of mutations that make the difference between “darwinism” and mutationism. If natural selection merely acts to fix rare mutations of large effect then it can’t be called creative or called the engine of adaptation. Is that it?