In grad school I wound up hanging around with John Pepper (yeah, Dr. Pepper) a good bit. I think I disagreed with him more than I agreed with him, sometimes to the point of exasperation, but conversations with him were never boring.

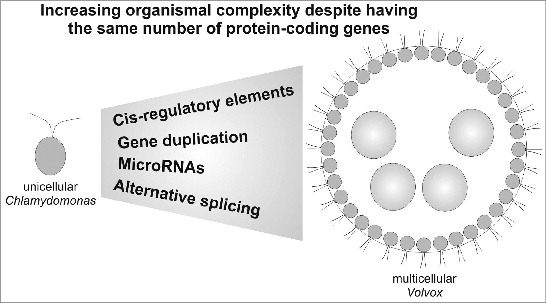
One of John’s most annoying refrains was “is it an organism?” I was studying (and still study) a group of algae for which this question can be genuinely confusing. Most people would say a Chlamydomonas cell is a single-celled organism, and most would agree that Volvox is a multicellular organism, but what about the four-celled species Tetrabaena? A four-celled organism or a collection of four single-celled organisms? What about an undifferentiated colony of 32 cells, such as Eudorina? Or Pleodorina, which is around the same size but with two cell types? Somewhere between a unicellular ancestor and Volvox, a new kind of individual emerged. Among the extant species*, where do we draw the line between organisms and groups of organisms, or can we (or should we) draw a line at all?